|

|
|
|
7.2 Le disque embryonnaire tridermique (3ème semaine)
|
|
|
Formation des feuillets primordiaux
|
|
|
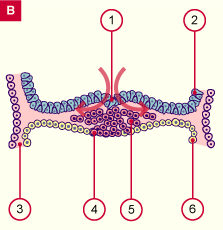
| Fig. 3 - Ligne primitive vers 18 jours vue dorsale |
|
Légende |
1
2
3
4
5
6
7
8
9
NB |
sillon primitif
dépression primitive
noeud primitif
membrane oropharyngée
aire cardiaque
bord sectionné de l'amnios
mésoderme
endoderme
future membrane cloacale
1+2+3=ligne primitive |
|
|
|
Fig. 3
Disque embryonnaire vu par sa face supérieure (dorsale) d'aspect ovoide, avec en haut la région céphalique, et en bas la région caudale.
|
L'épiblaste est à l'origine des trois couches cellulaires de l'embryon tridermique:
- l'ectoblaste
- le mésoblaste
- l'entoblaste définitif
|
|
|
|
|
|
Pour en savoir plus
|
|
Les molécules d'adhésion cellulaire ou CAM (Cell Adhesion Molecules)
Dès le début de la gastrulation les cellules épiblastiques produisent de l'acide hyaluronique, qui s'accumule dans l'espace intercellulaire entre l'épiblaste et l'hypoblaste. L'acide hyaluronique est fréquemment associé à la migration cellulaire. Cette molécule peut lier massivement l'eau (jusqu'à 1000 fois son propre poids !) et jouerait un rôle antiagrégant au niveau des cellules mésoblastiques. Toutefois la présence de l'acide hyaluronique ne suffit pas à expliquer la migration depuis la ligne primitive. Chez tous les embryons vertébrés cette dernière serait en outre liée à la présence de fibronectine associée à la lame basale sous l'épiblaste. La fibronectine (10) est une glycoprotéine extracellulaire.
Les intégrines
Les intégrines font partie des 4 familles de molécules d'adhérence CAM (Cell Adhesion Molecules): intégrines, cadhérines, sélectines, super famille des immunoglobulines. Elles assurent la jonction et la reconnaissance entre 2 cellules ou entre une cellule et la matrice extracellulaire. Leur mode de liaison est homophilique (molécules de même type) ou hétérophilique (molécules de type différent). Il s'agit de glycoprotéines transmembranaires assurant la cohésion (agrégation) et la migration cellulaire (responsable de l'organisation des cellules en tissus et des tissus en organes). Les intégrines forment des contacts avec les filaments de collagène dans la membrane basale et la matrice extracellulaire. La laminine est un ligand (molécule capable de s'attacher à un récepteur transmembranaire) de la membrane basale, alors que la fibronectine est un
ligand de la substance intercellulaire du mésenchyme et partant un support de la migration cellulaire.
|
|
|
|
|
|
|