|

|
|
|
|
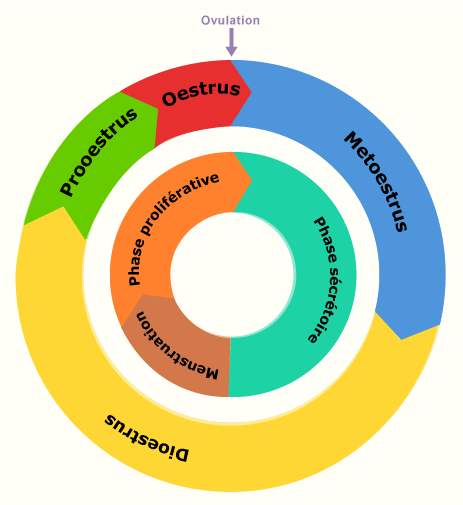
Le cycle féminin (cycle oestral) peut être divisé en quatre phases: le prooestrus (avant les chaleurs), l'oestrus (les chaleurs), le métoestrus (après les chaleurs) et le dioestrus (entre les chaleurs). Selon les espèces, une phase de repos ovarienne se produit et est appelée anoestrus,
|
|
|
| Fig. 10 - Vue comparative du cycle |
|
Légende |
|
|
Fig. 10
Comparaison des différentes phases du cycle chez la femme et chez les mammifères domestiques.
|
|
Le signe distinctif le plus caractéristique du premier jour du cycle chez la femme est l’apparition des règles. En principe, toutefois, d'autres événements récurrents peuvent tout aussi bien être utilisés pour définir le début du cycle.
Ainsi, chez les animaux domestiques, il existe différentes possibilités pour définir le premier jour du cycle. Les symptômes de chaleurs bien reconnaissables depuis l'extérieur pourraient être un des critères utilisés. Malheureusement, il n'existe pas de corrélation sans équivoque entre le début des chaleurs et un événement précis du cycle ovarien.
Lors de la représentation cyclique, l'ovulation est désignée comme premier jour du cycle (temps 0) et ceci indépendamment de symptômes externes.
L'ovulation est sans aucun doute un événement approprié, précis et bien défini. Malheureusement il ne peut pas être déterminé sans hésitation depuis l'extérieur.
En raison de ces différentes possibilités, il est absolument nécessaire de spécifier la définition du début du cycle choisie. Sauf indication contraire, l'ovulation sera considérée dans le texte suivant comme le début du cycle.
|
|
|
|
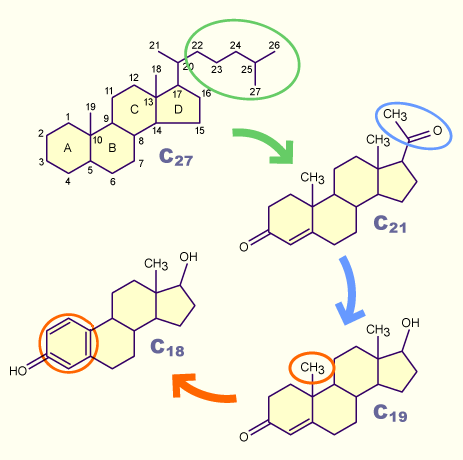
Dans le prooestrus, également appelé phase de maturation folliculaire, le niveau hormonal est successivement régi par les gonadotrophines (FSH et LH). Les niveaux élevés de progestérone de la phase lutéale précédente diminuent. Etant donné que les cellules de la granulosa expriment les récepteurs à la FSH, l'augmentation des taux de FSH stimule la maturation de follicules pré-antraux et antraux.
Le taux élevé de FSH est associé d'une part à une réactivité augmentée des follicules aux hormones gonadotropes, en particulier la FSH, et d'autre part à une augmentation de la production d'œstrogènes. La biosynthèse de ces hormones stéroïdiennes est initiée dans les cellules de la thèque interne à partir du cholestérol (C27) et passe par la formation de gestagènes (C21) puis d'androgènes (C19).
La transformation des androgènes (C19) en œstrogènes (C18) nécessite une transformation d'un anneau de cyclohexane en cycle aromatique par le biais d'une oxydation et de la perte d'un second groupement méthyle. Ces étapes biochimiques se déroulent exclusivement dans les cellules de la granulosa de l'épithélium folliculaire.
|
|
|
| Fig. 11 - Stéroïdogenèse |
|
Légende |
|
|
Fig. 11
La biosynthèse d'hormones stéroïdiennes est initiée à partir du cholestérol (C21). Après séparation de la chaîne latérale polaire, le groupe de progestatifs (C21) se forme. L'élimination de deux autres atomes de carbone permet la formation des androgènes (C19). La formation du groupe des oestrogènes est atteinte grâce à la suppression d'un autre groupe méthyle et la transformation d'un anneau de cyclohexane en cycle aromatique.
La stéroïdogenèse ne se réalise que dans cette direction et n'est donc pas réversible.
|
|
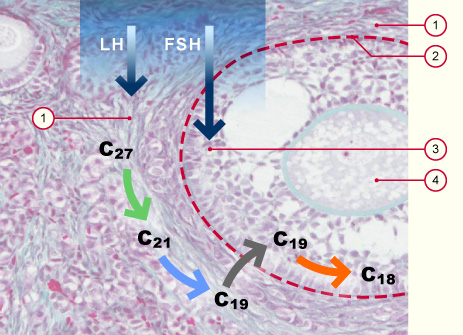
La thèque interne étant perfusée grâce à un riche réseau de capillaires, ses cellules peuvent absorber le cholestérol sanguin. L'épithélium folliculaire est lui avasculaire et n'a donc pas accès au cholestérol sanguin.
L'influence de la LH se fait sur les cellules de la thèque, alors que les cellules épithéliales folliculaires expriment les récepteurs de la FSH.
|
|
|
| Fig. 12 - Aromatase |
|
Légende |
1
2
3
4
|
Cellules de la thèque
Lame basale entre la thèque (1) et l'épithélium folliculaire (3)
Epithélium folliculaire
Ovocyte
|
|
|
|
|
|
Fig. 12
Absorption du cholestérol (C27) depuis le sang et métabolisation ultérieure en C21 et C19 par les cellules de la thèque sous l'influence de la LH.
Absorption d'androgène (C19) par l'épithélium folliculaire et conversion en œstrogène sous l'influence de la FSH.
|
|
L'aromatase revêt ainsi une importance majeure pour la détermination des caractéristiques sexuelles phénotypiques. Elle transforme les androgènes virilisants synthétisés par la thèque en œstrogènes féminisants. La biosynthèse des stéroïdes sexuels ne s'effectue que dans un sens, de sorte que les œstrogènes ne peuvent pas être reconvertis en androgènes qui eux ne pourront pas être convertis en progestatifs.
Au cours du cycle sexuel, la diminution du rapport progestérone-œstrogène entraîne des modifications typiques au niveau du tractus génital et du comportement
|
|
|
|
Au début de l'oestrus, les taux élevés d'oestrogène sont responsables de nombreux changements internes et externes.
Ils augmentent l'irrigation sanguine et l'oedématisation des muqueuses du tractus génital, élargissent le canal cervical, augmentent la viscosité de la glaire cervicale et entraînent une augmentation des cellules ciliées de l'ampoule de la trompe utérine.
Les œstrogènes augmentent également la contractilité du myomètre et sa réponse à l'ocytocine.
Extérieurement, les œstrogènes sont responsables des symptômes de chaleur spécifiques à l'espèce, d'une oedématisation de la vulve et du comportement typique lors de chaleurs (disposition à s'accoupler).
Le mécanisme de rétrocontrôle positif entre œstrogène et gonadolibérine (GnRH; Gonadotropin Releasing Hormon) entraîne une nouvelle augmentation de la FSH et qui est suivie d' une augmentation à court terme de la libération de LH. Ce pic LH déclenche la poursuite de la deuxième division méiotique (division équationnelle). Une nouvelle interruption de la méiose a lieu en métaphase II.
Le pic LH permet également une maturation globale du follicule et permet une ovulation soit unique chez les espèces unipares ou multiple chez les espèces multipares. La paroi folliculaire s'amincit et la cavité folliculaire s'ouvre. Chez certaines espèces, telles que le chat et le lapin, le stimulus mécanique de la copulation est nécessaire en plus du pic de LH pour déclencher l'ovulation (ovulation induite).
L'ovulation est le résultat d'interactions hormonales, enzymatiques et mécaniques complexes. Il faut toutefois noter que tous ces mécanismes ne sont pas élucidés dans tous les détails. Par le biais du flux sanguin accru au niveau du follicule, la paroi folliculaire est hyperémiée, une vasodilatation est observée et il se forme une saillie conique du follicule à la surface de l'ovaire (Stigma follicullaire = site de l'ovulation).
A ce moment, sous l'influence d'hormones spécifiques à l'espèce animale, le débit sanguin dans le follicule se réduit. Il en résulte une anémie locale dans la zone du stigma conduisant à la lyse des cellules et du stroma. Cela permet l'expulsion du Cumulus oophorus contenant l'ovule. Contrairement à d'anciennes hypothèses qui supposaient une augmentation de la pression intrafolliculaire responsable de l'ovulation, des mesures de pression ont montré que la pression dans le follicule chutait peu avant l'ovulation.
L’infundibulum tubaire ou pavillon capte le Cumulus oophorus et le transporte dans l'ampoule. Le liquide folliculaire expulsé devient gélatineux et entoure l'ovule et les cellules du cumulus. Les ovules, à l'exception des ovules de la chienne (voir chapitre cycle sexuel, chien), ne restent fécondables que quelques heures.
Selon l'espèce, les symptômes de chaleur régressent déjà avant l'ovulation ou y survivent.
|
|
|
|
Après l'ovulation, la cavité résultant de l'écoulement du liquide folliculaire avec le Cumulus oophorus collabe et s'emplit d'un exsudat séro-fibrineux et de sang. Il se forme le Corpus hemorrhagicum. Il se transformera par la suite en Corpus luteum (corps jaune mature), Corpus rubrum (corps jaune en régression) et Corpus albicans (cicatrice de tissu conjonctif)
Les capillaires sanguins provenant de la paroi folliculaire collabée prolifèrent à travers la membrane basale dans l'épithélium folliculaire restant. Dans le même temps, on observera une lutéinisation des cellules épithéliales folliculaires en lutéocytes granulaires (granulosoluteocyti) et des cellules stromales en lutéocytes thécaux (thecoluteocyti). En conséquence, cette phase est également appelée phase de formation du corps jaune et correspond au metoestrus.
En raison de l'augmentation de la progestérone, les changements régis par les œstrogènes vont régresser. D'autre part, les glandes utérines seront stimulées pour sécréter le lait utérin. Celui-ci est la première source nutritive du foetus (histiotrophie).
Durant le metoestrus l'ovule ou les ovules (animaux multipares) se trouvent dans la trompe. La fécondation se fera à ce niveau.
La transition entre le metoestrus et le dioestrus se fait de manière progressive.
|
|
|
|
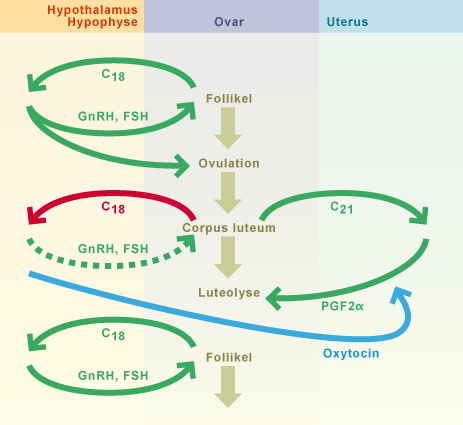
Avec la transformation ultérieure des cellules folliculaires en lutéocytes, le corps jaune fonctionnellement actif (ou les corps jaunes fonctionnellement actifs) se forme. Un corps jaune en fleur perdure durant tout le dioestrus. La croissance et la durée de vie du corps jaune dépendent de facteurs lutéotropes spécifiques à l'espèce. La formation des lutéocytes s'accompagne également d'une augmentation de la production de progestérone, qui elle définit le dioestrus
Les progestatifs inhibent la libération de GnRH (Gonadotropin Releasing Hormon, gonadolibérine). La sécrétion de GnRH n'est, cependant, jamais complètement bloquée dans l'hypothalamus et des petites quantités de FSH et de LH seront libérées permettant ainsi au processus de maturation folliculaire de se poursuivre. Il n'y aura cependant pas d'ovulation sous dominance de progestérone sauf chez la jument (voir le chapitre cheval). Au niveau de l'utérus, la progestérone stimule les glandes utérines à produire le lait utérin (mucus nutritif vital pour l’embryon durant les premières semaines de vie). Sous l'effet de la progestérone, la glaire cervicale a tendance à devenir plus liquide et sa production se réduit.
Si l'endomètre ne reçoit aucun signal de la part d'un embryon ou, en d'autres termes, si l'organisme maternel ne détecte aucune gestation, la lutéolyse commence et le processus cyclique se poursuit avec le prooestrus. Chez toutes les espèces animales, la prostaglandine F2α qui se forme dans la paroi utérine est l’élément déclencheur de la lutéolyse (résorption du corps jaune).
L'échange veino-artériel entre la veine ovarienne (Vena ovarica) et l'artère ovarienne (Arteria ovarica) permet d'atteindre localement dans le corps jaune des taux élevés de prostaglandines.
|
|
|
| Fig. 13 - Hormone |
|
Légende |
|
|
Fig. 13
La régression du corps jaune entraîne une chute du taux de progestérone. Son effet inhibiteur sur le système hypothalamus / hypophyse (suppression du rétrocontrôle négatif) diminue et la libération de GnRH augmente à nouveau. Subséquemment, les taux de FSH augmentent et ainsi entraîneront une maturation folliculaire.
Les oestrogènes produits par les follicules provoqueront à leur tour les caractéristiques du prooestrus.
|
|
|

