|

|
|
|
|
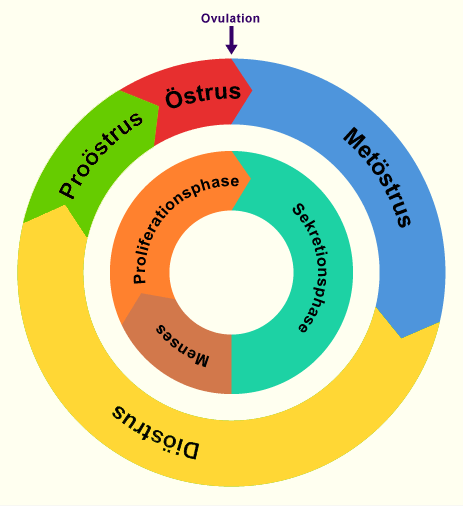
Der weibliche Zyklus kann in vier Phasen eingeteilt werden: Proöstrus (Vorbrunst), Östrus (Brunst), Metöstrus (Nachbrunst), Diöstrus (Zwischenbrunst). Je nach Spezies kommt eine Ovarruhephase vor, die als Anöstrus bezeichnet wird.
|
|
|
| Abb. 10 - Zyklus vergleichend |
|
Legende |
|
|
Abb. 10
Gegenüberstellung der Zyklusphase beim Menschen und bei den Haussäugetieren.
|
|
Beim Menschen wird der Beginn der Menstruation als das auffälligste Merkmal als 1. Zyklustag bezeichnet. Grundsätzlich können aber andere wiederkehrende Ereignisse ebenso gut als Beginn des Zyklus herangezogen werden. So bestehen bei den Haustieren verschiedene Möglichkeiten, den 1. Zyklustag zu definieren. Als äusserlich gut erkennbares Merkmal bietet sich bei den Haustieren der Beginn der Brunstsymptome für die Bestimmung des Zyklus an. Allerdings ist der Brunstbeginn in Bezug auf den ovariallen Zyklus nicht mit einem eindeutigen Ereignis korreliert. Bei der Darstellung des zyklischen Geschehens wird, unabhängig von äusserlichen Symptomen, die Ovulation als 1. Zyklustag (Zeitpunkt 0) bezeichnet. Als Startpunkt eines Zyklus ist die Ovulation zweifellos ein geeignetes, präzise definiertes Ereignis. Allerdings lässt sie sich nicht ohne weiteres von aussen feststellen. Aufgrund dieser unterschiedlichen Möglichkeiten ist es zwingend notwendig, bei Angaben zu Zyklustagen die gewählte Festlegung des Zyklusbeginns zu präzisieren. Sofern nicht anders ausgewiesen, gilt im folgenden Text jeweils die Ovulation als Zyklusbeginn.
|
|
|
|
Im Proöstrus, der auch als Follikelreifungsphase bezeichnet wird, verschiebt sich der Hormonspiegel zugunsten der Gonadotropine (FSH und LH). Die hohen Progesteronwerte aus der vorhergehenden Lutealphase fallen ab. Der Anstieg des FSH-Spiegels fördert die Reifung der präantralen und antralen Follikel, da deren Granulosazellen FSH-Rezeptoren exprimieren.
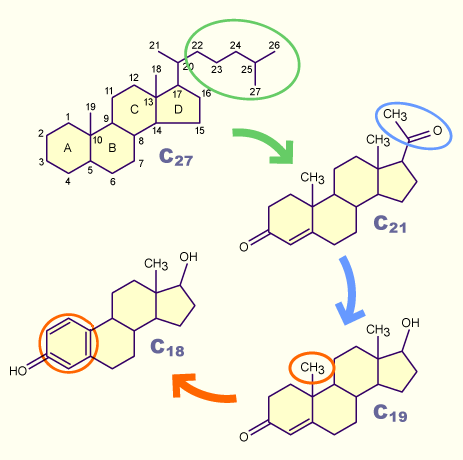
Der hohe FSH-Spiegel geht einerseits mit einer erhöhten Ansprechbarkeit der Follikel auf die gonadotropen Hormone, insbesondere FSH, einher und führt andererseits zur vermehrten Produktion von Östrogenen. Die Bildung dieser Steroidhormone geht aus vom Cholesterin (C27), welches in den Zellen der Theka interna über Gestagene (C21) zu Androgenen (C19) metabolisiert wird.
Die Umwandlung der Androgene (C19) in Östrogene (C18) erfordert die Abspaltung einer weiteren Methylgruppe und die Aromatisierung eines Zyklohexanrings, biochemische Etappen, welche ausschliesslich in den Granulosazellen des Follikelepithels erfolgen.
|
|
|
| Abb. 11 - Steroidogenese |
|
Legende |
|
|
Abb. 11
Die Bildung der Sexualsteroide geht aus vom Cholesterin (C21). Nach Abtrennung der polaren Seitenkette entsteht die Gruppe der Gestagene (C21). Aus diesen werden durch Entfernung von zwei weiteren C-Atomen die Androgene (C19) gebildet. Durch Entfernung einer weiteren Methylgruppe und Aromatisierung des Zyklohexanrings A wird die Gruppe der Östrogene erreicht.
Die Steroidogenese verläuft nur in dieser Richtung und ist damit nicht umkehrbar.
|
|
Die Theka interna ist durchblutet, so dass die Thekazellen Blutcholesterin aufnehmen können. Das Follikelepithel hingegen ist avaskulär und hat damit keinen Zugang zum Blutcholesterin.
Die Thekazellen stehen unter dem Einfluss von LH, die Follikelepithelzellen exprimieren hingegen FSH-Rezeptoren.
|
|
|
| Abb. 12 - Aromatase |
|
Legende |
1
2
3
4
|
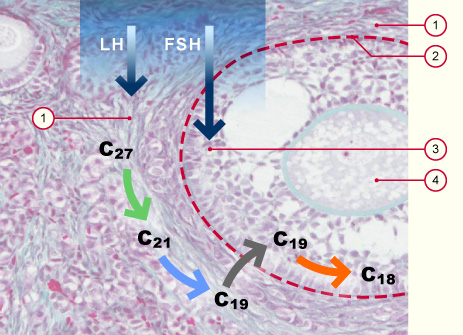
Theka-Zellen
Basalmembran zwischen Theka folliculi (1) und Follikelepithel (3)
Follikelepithel
Oocyte
|
|
|
|
|
|
Abb. 12
Aufnahme von Cholesterin (C27) aus dem Blut und weitere Metabolisierung zu C21 und C19 durch die Theka-Zellen unter dem Einfluss von LH.
Aufnahme der Androgene (C19) durch das Follikelepithel und Umwandlung zu Östrogenen unter dem Einfluss von FSH.
|
|
Damit kommt der Aromatase auch eine wesentliche Bedeutung für phänotypische Geschlechtsmerkmale zu, transformiert sie doch die virilisierenden Androgene in feminisierende Östrogene. Die Biosynthese der Sexualsteroide läuft nur in einer Richtung ab, so dass die Östrogene nicht in Androgene, und diese nicht in Gestagene zurückverwandelt werden können.
Das sinkende Progesteron-Östrogen-Verhältnis zieht im Laufe des Sexualzyklus typische Veränderungen am Genitaltrakt und im Verhalten nach sich.
|
|
|
|
Am Anfang des Östrus dominieren die hohen Östrogenspiegel das Geschehen: sie verstärken die Durchblutung und Ödematisierung der Schleimhäute im Genitaltrakt, erweitern den Cervikalkanal, erhöhen die Viskosität des Cervixschleimes und führen zu einer Vermehrung der Zilienzellen in der Ampulla des Eileiters. Östrogene erhöhen ausserdem die Kontraktilität des Myometriums und dessen Ansprechbarkeit auf Oxytocin. Äusserlich bewirken die Östrogene speziesspezifische Brunstsymptome, eine Ödematisierung der Vulva und das brunsttypische Verhalten (Paarungsbereitschaft).
Der positive Feedbackmechanismus zwischen Östrogen und dem Gonadotropin Releasing Hormon (GnRH) führt zu einem weiteren FSH-Anstieg und kurz darauf zu einer kurzzeitig erhöhten Ausschüttung von LH. Dieser LH-Peak löst die Fortsetzung der Reduktionsteilung beziehungsweise die Äquationsteilung aus. In der Metaphase II kommt es jedoch zu einem weiteren Unterbruch der Meiose.
Der LH-Peak lässt auch den Follikel als Ganzes ausreifen und ermöglicht die Ovulation (unipare Spezies) beziehungsweise die Ovulationen (multipare Spezies), indem die Follikelwand ausdünnt und die Follikelhöhle eröffnet wird. In einigen Spezies, zum Beispiel bei der Katze und beim Kaninchen, braucht es zustäzlich zum LH-Peak den mechanischen Stimulus der Kopulation zur Auslösung der Ovulation (induzierte Ovulation).
Die Ovulation ist das Resultat komplexer hormoneller, enzymatischer und mechanischer Wechselwirkungen, wobei der genaue Mechanismus nicht vollständig geklärt ist. Durch die verstärkte Durchblutung des Follikels wird die Follikelwand hyperämisiert, erfährt eine Vasodilatation, und es bildet sich eine Vorwölbung des Follikels an der Ovaroberfläche (Stigma = Ovulationsstelle). Durch tierartlich spezifische hormonelle Einflüsse wird nun der Blutfluss im Follikel vermindert. Die daraus resultierende lokale Anämie im Stigmabereich führt zur Lyse von Zellen und Grundsubstanz. Dadurch kann der Cumulus oophorus, der die Eizelle beinhaltet, aus dem Follikel austreten. Entgegen früherer Annahmen, die einen erhöhten intrafollikulären Druck für die Ovulation verantwortlich machten, konnte mit Druckmessungen nachgewiesen werden, dass der Druck im Follikel kurz vor der Ovulation absinkt.
Das Infundibulum nimmt den Cumulus oophorus auf und transportiert ihn in die Eileiterampulle. Die ausgetretene Follikelflüssigkeit wird gelartig und umgibt die Eizelle und die Cumuluszellen. Die Eizellen bleiben, mit Ausnahme der Eizellen der Hündin (siehe Kapitel Sexualzyklus, Hund), nur für Stunden befruchtungsfähig.
Je nach Spezies gehen die Brunstsymtpome bereits vor der Ovulation zurück oder aber sie überdauern diese.
|
|
|
|
Die bei der Ovulation durch das Ausfliessen von Follikelflüssigkeit mit Cumulus oophorus entstandene Höhle kollabiert und füllt sich mit Gewebeflüssigkeit und Blut, es bildet sich das Corpus hemorrhagicum. Dieses wird sich im Laufe seiner Entwicklung in das Corpus luteum (Gelbkörper in Blüte), Corpus rubrum (Gelbkörper in Rückbildung) und Corpus albicans (alte, bindegewebige Gelbkörpernarbe) umwandeln.
Die Blutkapillaren aus der kollabierten Follikelwand sprossen durch die Basalmembran in das verbliebene Follikelepithel ein. Gleichzeitig erfolgt die Luteinisierung der Follikelepithelzellen zu Granulosa-Luteinzellen, jene der Stromazellen zu Theka-Luteinzellen. Deshalb wird diese Phase auch als Gelbkörperanbildungsphase bezeichnet. Sie entspricht dem Metöstrus.
Durch den Progesteronanstieg bilden sich einerseits die durch Östrogeneinfluss entstandenen Veränderungen zurück, andererseits werden die Uterindrüsen zur Sekretion von Uterinmilch angeregt. Diese histiotrophe Ernährung stellt die erste Versorgung des Keimlings dar.
Im Metöstrus befindet sich die Eizelle (beziehungsweise befinden sich die Eizellen multiparer Tiere) im Eileiter, wo gegebenenfalls eine Befruchtung stattfindet).
Der Übergang zwischen Metöstrus und Diöstrus ist fliessend.
|
|
|
|
Mit der weiteren Umwandlung der Follikel- in Luteinzellen entsteht das funktionell aktive Corpus luteum (beziehungsweise die funktionell aktiven Corpora lutea). Ein Gelbkörper in Blüte bleibt für die Dauer des Diöstrus bestehen. Anbildung und Lebensdauer des Gelbkörpers hängen von speziesspezifischen luteotropen Faktoren ab. Mit der Ausbildung der Luteinzellen geht auch eine zunehmende Progesteronproduktion einher, die den Diöstrus definiert.
Gestagene hemmen die Ausschüttung von GnRH. Die Sekretion von GnRH wird jedoch nie vollständig unterdrückt, so dass auch im Diöstrus geringe Mengen an FSH und LH ausgeschüttet werden. Deshalb läuft der Prozess der Follikelreifung weiter, es kommt jedoch normalerweise zu keinen Ovulationen unter Progesterondominanz mit Ausnahme der Stute (siehe Kapitel Pferd). Im Uterus veranlasst Progesteron die Uterindrüsen zur Sekretion der Uterinmilch, während die Beschaffenheit des Cervikalschleim eher wässrig und dessen Produktion vermindert wird.
Erhält das Endometrium keine Signale eines Embryos oder, anders gesagt, erkennt der mütterliche Organismus keine Trächtigkeit, so setzt die Luteolyse ein und damit setzt sich das zyklische Geschehen mit dem Proöstrus fort. Auslöser der Luteolyse ist bei allen Tierarten das Prostaglandin F2α, das in der Uteruswand gebildet wird. Über einen veno-arteriösen Austauschvorgang zwischen Vena ovarica und Arteria ovarica werden lokal hohe Prostaglandinspiegel im Gelbkörper erreicht.
|
|
|
| Abb. 13 - Hormone |
|
Legende |
|
|
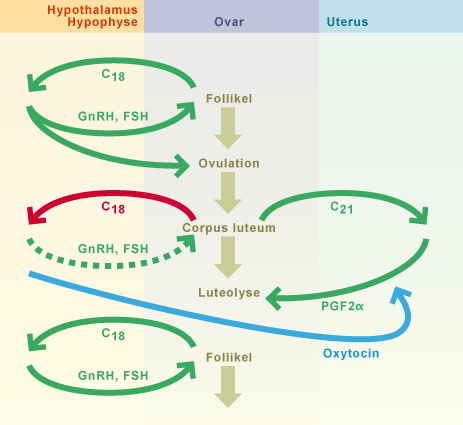
Abb. 13
Die Rückbildung des Gelbkörpers führt zu einem Abfallen des Progesteronspiegels. Dadurch entfällt dessen hemmende Wirkung auf das HT/HVL-System, und die Ausschüttung von GnRH nimmt wieder zu. Der FSH-Spiegel steigt als Folge davon an und bewirkt die Follikelreifung.
Die von den Follikeln gebildeten Östrogene rufen ihrerseits die Merkmale des Proöstrus hervor.
|
|
|

